|
||

|
肝脏被结缔组织被膜包绕,该被膜称为格利森被膜(Glisson被膜)。 | |
|
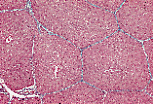
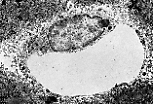
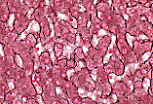
猪的肝脏可见界限清晰的独立小叶。
每个多面体形的小叶被薄层结缔组织分隔包绕。 |
|
|
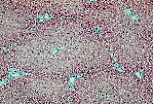
人类肝脏的小叶无结缔组织分隔;可通过小叶角部存在的结缔组织-血管间隙(即门管区)来判断小叶的边界。 | |
|
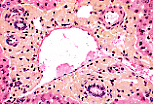
门管区是一种结缔组织结构,包含门静脉分支、肝动脉分支、 毛细淋巴管 和胆管。 | |
|
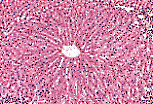
每个小叶由肝细胞组成的肝细胞索(雷马克索,travées de Remak)构成;肝细胞索围绕中央静脉(小叶中央静脉)呈放射状排列。
来自门静脉分支的血液在雷马克索之间的血窦中流动,血窦汇流至中央静脉,再由中央静脉将血液输送至肝静脉。 |
|
|
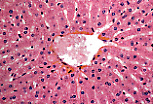
中央静脉接收流经肝实质血窦的血液。
因此,肝血窦构成了一个 独特的毛细血管网 ,连接于两条静脉之间。 |
|
|
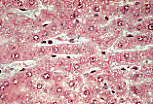
雷马克索由单层上皮样肝细胞板构成,肝细胞板可局部吻合;
血窦占据肝细胞索之间的间隙。 |
|
|
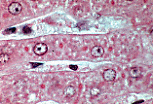
肝细胞为多面体细胞,细胞核染色浅,含1-2个大核仁;细胞质呈颗粒状。
嗜碱性最强的区域对应粗面内质网聚集区,染色浅的区域则为糖原或脂质包涵体。
每个肝细胞有两个血管极(或血管面)朝向血窦,以及肝细胞间的接触面(称为胆小管极); 肝细胞通过质膜内陷形成胆小管,肝细胞产生的胆汁排入胆小管中。 |
|
|
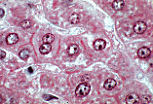
肝细胞常可见双核,或细胞核体积大于平均水平(多倍体细胞)。 正常成年肝脏中核分裂象极少见,因为正常情况下肝细胞的更新速度极慢。 | |

|
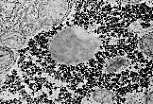
PAS染色证实肝细胞质富含糖原。 | |
|
透射电镜(MET)下,糖原表现为电子致密颗粒,呈玫瑰花结状聚集;也可识别出脂质包涵体。 | |
|
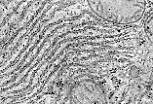
肝细胞内粗面内质网高度发达,由平行的扁平囊泡聚集而成。
光面内质网较不明显,呈管状网络,与粗面内质网相连。 |
|
|
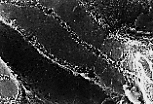
扫描电镜(MEB)下观察肝板断面,可见肝细胞的血窦面布满微绒毛,
胆小管面较光滑,有胆小管穿行且胆小管内也布满微绒毛。
肝血窦 可通过不连续的管壁和大小不等的窗孔识别; 血窦内皮直接贴附于肝细胞血管面的微绒毛上。 |
|
|
血窦内皮与肝细胞微绒毛之间存在狭小的间隙,即狄氏间隙(Disse间隙);该间隙仅能在透射电镜下观察到。 | |

|
该视野清晰显示肝血窦内皮的窗孔,以及狄氏间隙;肝细胞微绒毛浸泡于狄氏间隙的液体中。 | |
|
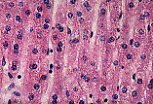
光镜(MO)下,固定良好的标本中无法观察到狄氏间隙; 可通过细长的细胞核识别内皮细胞,内皮细胞紧密贴合肝细胞的血管面。 | |
|
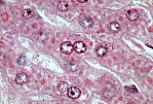
狄氏间隙内存在纤细的网状纤维网,可通过银染法显示。 | |
|
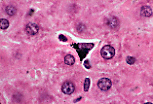
除内皮细胞外,血窦内还可见体积更大的细胞,细胞核形态不规则、有时弯曲,即枯否细胞(Kupffer细胞)。 | |
|
给大鼠静脉注射墨汁后,其肝脏标本可显示枯否细胞的吞噬功能: 墨汁颗粒出现在枯否细胞的胞质中,而不在内皮细胞内。 | |
|
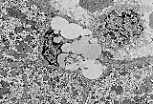
该视野显示人肝中的伊东细胞(Ito细胞);
该细胞位于狄氏间隙内,可通过弯曲的细胞核和脂质包涵体识别。
伊东细胞的功能之一是储存维生素A。 |
|
|
||
| 胆小管 | ||

|
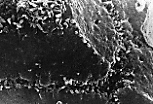
扫描电镜下可见肝内胆管的起始部——胆小管;
胆小管是细长的管道,由相邻肝细胞胆小管面的线性质膜凹槽相互贴合形成。
因此,胆小管无自身管壁,仅由肝细胞的细胞膜界定。 胆小管在肝板内形成连续、吻合的细胞周网络,不与血窦相通。 |
|
|
胆小管凹槽内的微绒毛增加了肝细胞的胆汁分泌表面积。 | |
|
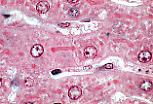
光镜高倍镜下可分辨胆小管,其边缘被伊红染色而强化。
胆小管总是位于肝细胞之间,呈管状或线状。 |
|
|
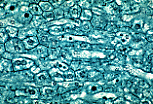
银染法可显示胆小管,此时胆小管呈六角形网格状。 | |
|
胆小管两侧的连接复合体保证了完美的密封性,使胆汁不与血窦内的血液接触。 | |
| 闰管 - 赫林管(Hering管) | ||
|
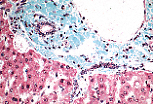
胆小管将肝细胞分泌的胆汁输送至门管区(与血液流动方向相反)。
接近门管区时,胆小管开口于短的、有独立管壁的管道; 该管道先由扁平细胞构成,后变为立方细胞,称为闰管(cholangioles)。 当闰管进入门管区时,有时称为赫林管(Hering管)。 |
|
|
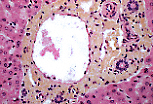
赫林管位于门管区周边,可通过狭窄的管腔和单层立方上皮识别。 | |
| 胆管 | ||
|
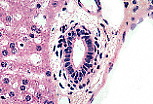
赫林管汇合成管径更大的胆管,胆管衬覆单层柱状上皮, 上皮外有少量同心圆排列的结缔组织支持。 | |
|
管径较大的胆管中,柱状上皮细胞的顶端会形成纹状缘。
上皮外有结缔组织-肌层包绕;胆管越粗,平滑肌细胞越丰富。 该管壁延续至肝外胆管(肝管和胆总管)。 |
|